|






 |  |

| Medicina | Medicina veterinara | Muzica | Psihologie | Retete | Sport |
Medicina
|
|
Qdidactic » sanatate & sport » medicina Mecanismele de actiune a hormonilor |
Mecanismele de actiune a hormonilor
Mecanismele de actiune a hormonilor
1. Receptorii hormonali si activarea lor
Hormonii se combina cu receptorii hormonali de pe suprafata membranelor celulare sau din interiorul celulelor. Combinatia dintre hormon si receptor declanseaza in celula o cascada de reactii. Toti sau aproape toti receptorii hormonali sunt proteine mari si fiecare receptor este aproape totdeauna foarte specific pentru un anumit hormon.
Receptorii in stare nelegata sunt inactivi. Exista cai diferite de activare in functie de tipul receptorului. exista posibilitatea unui efect direct al unor receptori hormonali de a modifica permeabilitatea membranei celulare sau mai important doua mecanisme generale: (1) activarea sistemului AMPc din celule, care apoi activeaza alte functii intracelulare sau (2) prin activarea genelor celulare, care determina formarea unor proteine intracelulare si care la randul lor produc activarea specifica a unor functii celulare
Principala cale de transport a hormonilor este sangele circulant, in care hormonul se afla sub forma libera sau legati de un transportor mai mult sau mai putin specific. In general hormonii proterici circula liberi in sange, in timp ce hormonii tiroidieni circula legati de molecule proteice. Este important de retinut ca forma libera este forma activa.
Semi-viata unui hormon in circulatie depinde de proportia dintre forma legata si cea libera. Tiroxina care circula in 99% in forma legata, are semiviata lunga de 6 zile iar aldosteronul ca se leaga in proportie de 60% are a semiviasa de 15 minute.
Carausii plasmatici nu sunt inerti, ei pot elibera hormonul liber, activ, constituind o rezerva circulanta de hormoni. Drogurile care intra in competitie cu transportorii specifici pot modifica durata efectului unui hormon. De pilda, aspirina intra in comperitie cu transportorii proteici ai hormonilor tiroidieni, prelungind efectul acestora din urma, fapt nu lipsit de importanta in situatiile in care hormonii tiroidieni sunt excitati in exces.
Majoritatea hormonilor sunt prezenti in sange in cantitati extrem de mici, unele in concentratii de o milionime dintr-un miligram (1 picogram) pe mililitru.
In ultimii ani s-a dezvoltat o metoda extrem de sensibila de dozare a hormonilor, a precursorilor lor si a produsilor finali de metabolism, este metoda radioimunodifuzarii (RIA-radioimmunoassay)
1.1. Receptorii membranari de suprafata
Acesti receptori se combina specific cu un anumit hormon, pe care il recunoaste dupa conformatia sa spatiala specifica; semnalul mesagerului chimic de ordinul I (hormonul) este transferat de catre receptor in interiorul celulei cu ajutorul mesagerilor de ordinul II sau III.
Numarul receptorilor de suprafata (2000-10000/celula) este reglabil prin insusi nivelul circulant al hormonilor. In mod obisnuit cresterea concentratiei circulante de hormon activ induce reducerea numarului receptorilor pe suprafata celulei tinta; acesta este efectul "down-regulation". Efectele cresterii cronice a concentratiei hormonului in sange poat fi astfel contracarate cel putin partial prin acest mecanism de autoreglare la nivelul receptorului. In unele cazuri, excesul hormonal induce cresterea numarului receptorilor, urmata de amplificarea crescanda a raspunsului tinta, acesta fiind efectul de "up-regulation".
Anumite conditii metabolice pot determina variatii ale numarului de receptori; de pilda, obezitatea induce scaderea numarului de receptori pentru insulina (de aici toleranta scazuta la glucoza a diabeticilor obezi nedependenti de insulina). Unele droguri pot avea efecte asemanatoare, de exemplu sulfonil ureea induce o crestere a numarului receptorilor pentru insulinpa.
|
Numarul receptorilor poate fi afectat si prin mecanisme autoimune (anticorpi anti-receptor).
Majoritatea hormonilor peptidici si proteici actioneaza asupra unor receptori membranari de suprafata.
Exista doua sisteme principale de mediatori de ordinul II asociati hormonilor proteici: sistemul AMPc si sistemul inozitoltrifosfat-diacilglicerol (IP3-DAG).
Receptorul membranar are in structura sa trei componente: un situs de legare a hormonului; un situs de cuplare a complexului hormon-receptor cu efectorul; un efector care genereaza mesagerul secund prin intermediul caruia se realizeaza efectele intracelulare ale hormonului. (Fig.1)
Sistemul de cuplare al complexului hormon-receptor cu efectorul este rerprezentat de sistemul proteinelor G (numele provine de la capacitatea de a fixa nucleotide ciclice cu guanina).
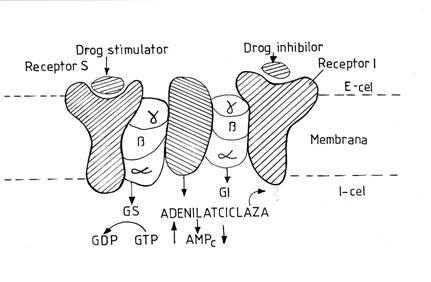
Fig,1. Receptorul membranar de suprafata-mesagerul secund, AMPc
Proteina G are o structura trimerica, adica trei subunitati proteice-alfa, beta, gamma.
Subunitatile beta si gama sunt comune pentru toate tipurile de proteine G. In functie de structura lantului alfa se disting mai multe tipuri de proteine G, notate Gs, Gi, Gq.
Proteinele G sunt situate pe fata interna a membranei celulare, fiind fixata prin intermediul subunitatii gamma in stratul bilipidic. Activarea proteinelor G se face prin legarea hormonului cu situsul receptor. In urma activarii, GDP-ul fixat pe subunitatea alfa este inlocuit cu o molecula guoanozin-trifosfat (GTP).
Aceasta substituire determina desprinderea subunitatii alfa de pe celelalte doua subunitati, interactionand cu sistemul efector reprezentat de o enzima membranara cu rol in generarea mesagerului secund. Subunitatea alfa a proteinei G are activitate GTP-azica, hidrolizand GTP in GDP in cateva secunde si producand revenirea la forma sa inactiva (alfa-GDP), care se reasociaza cu subunitatea beta si gamma. In consecinta se restabileste proteina G in forma sa de repaus, ce se disociaza de pe efctor.
Exista doua enzime-efector principale, implicate in mecanismul de actiune al hormonilor proteici: adenilatciclaza si fosfolipaza C.
Adenilatciclaza este activata de receptori hormonali cuplati cu proteinele Gs (stimulatoare) si inhibata de receptorii hormonali cuplati cu proteinele Gi (inhibitoare). Ea catalizeaza transformarea ATP-ului in AMPc, care reprezinta mesagerul secund. Acesta activeaza o enzima celulara- proteinkinaza A, care in forma activata catalizeaza fosforilarea unor proteine celulare cu rol de enzima care realizeaza efectele intracelulare ale hormonului. AMPc este degradat de o enzima specifica numita fosfodiesteraza. Prin intermediul AMPc actioneaza mai multi hormoni, cum ar fi hormonul antidiuretic, hormonii tropi hipofizari, parathormonul, glucagonul, adrenalina (prin intermediul beta-receptorilor adrenergici).
Fosfolipaza C este activata de receptori hormonali cuplati cu proteina Gq. Actioneaza asupra unui fosfolipid membranar-fosfatidil-inozitol-difasfat, pe care il scindeaza in inozitol-trifosfat (IP3) si diacilglicerol (DAG). IP3 difuzeaza in reticulul sarcoplasmatic unde functioneaza ca un eliberator de Ca2+ in citoplasma. Efectele intracelulare ale Ca2+ sunt mediate dupa o cuplare cu o proteina fixatoare de Ca2+ numita calmodulina, care poate fixa 4 ioni de Ca2+. Complexul Ca2+-calmodulina interactioneaza cu diferite enzime pe care le activeaza, presupunandu-se ca ar interveni in reglarea multora din functiile celulare. In mod cert intervine in diviziune celulara, mecanismele secretorii, contractia musculaturii netede. Dintre hormonii care asi exercita efectele prin eliberarea de Ca2+-intracelular amintin oxitocina, adrenalina, colecistochinina.
Diacilglicerolul ramane fixat pe membrana celulara si activeaza proteinkinaza C. Aceasta fosforileaza anumite proteine, schimbandu-le configuratia si totodata proprietatile functionale.
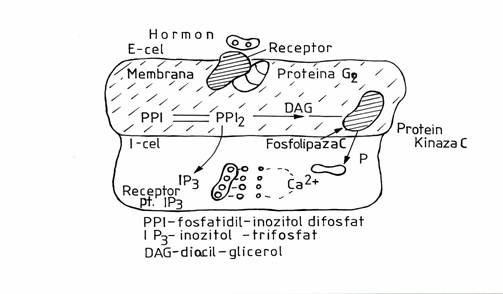
Fig.2-Mesagerul secund, Inozitoltrifosfat diacilgliceral
7. 2.1.2. Activarea genelor celulare
O a doua cale majora prin care unii hormoni actioneaza, in mod special hormonii secretati de cortexul adrenal, ovare si testicule, consta in determinarea sintezei de proteine in celulele tinta; unele dintre aceste proteine sunt enzime, care la randul lor activeaza alte functii celulare.
Secventa evenimentelor (pentru un hormon steroid) ar fi urmatoarea: hormonul patrunde in citoplasma celulara, unde se leaga cu un receptor proteic specific; hormonul combinat cu proteina receptor difuzeaza sau este transportat in nucleu; aceasta combinatie activeaza specific genele pentru a forma ARN mesager; ARN mesager difuzeaza in citoplasma promovand translatia la nivelul ribozomilor si sinteza de noi proteine.
Incetarea activitatii unui hormon se datoreaza unor mecanisme de metabolizare, urmata de excretia metabolitilor sau chiar a formei libere, active a hormonilor. Inactivarea enzimatica poate avea sediul chiar la nivelul receptorului sau in sedii tisulare multiple. Sediile cele mai importante ale inactivarii sunt ficatul si rinichii.
2. Reglarea secretiei hormonale
Capacitatea secretorie a glandelor endocrine este in mod normal mai mare decat necesarul hormonal al organismului.
Principalul mecanism de control al secretiei endocrine il constituie mecanismul de feed-back. Acesta functioneaza pe principiul cibernetic al sistemelor cu bucla de reactie. (Fig.3)
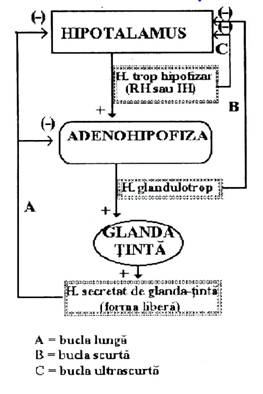
Fig.3- Reglarea secretiei prin mecanism de feed-back hormonal
Dupa modul in care functioneaza, se disting mai multe tipuri de bucle de feed-back:
1. Feed-back-ul hormonal- modificarea nivelului sanguin al unui hormon, modifica secretia endocrina a acelui hormon.
Dupa lungimea buclei reglatoare se disting:
feed-back scurt- in care modificarea nivelului sanguin al unui hormon influenteaza direct activitatea glandei care secreta acel hormon.
feed-back lung- in care modificarea nivelului sanguin al unui hormon influenteaza activitatea unei alte glande endocrine, care prin secretia ei influenteaza activitatea glandei care a secretat hormonul.
In acest sens intervine activitatea axului hipotalamo-hipofizar, care prin intermediul hormonilor tropi hipofizari, controleaza activitatea majoritatii glandelor endocrine.
2. Feed-back-ul nehormonal in care modificarea unor constante biologice dependente de nivelul hormonal, controleaza secretia glandelor endocrine respective. De exemplu, glicemia controleaza secretia de insulina; calcemia controleaza secretia de parathormon.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Documente online pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||