|






 |  |

| Medicina | Medicina veterinara | Muzica | Psihologie | Retete | Sport |
Medicina
|
|
Qdidactic » sanatate & sport » medicina Proprietatile functionale ale neuronilor |
Proprietatile functionale ale neuronilor
Proprietatile functionale ale neuronilor
Principalele proprietati functionale ale neuronilor sunt: generarea si conducerea impulsurilor nervoase.
Fenomenele electrice
Deoarece variatiile de potential in neuron si prelungirile sale sunt foarte reduse, a fost necesar sa se gaseasca tehnici suficient de fine pentru culegerea si amplificarea lor. Axonii neuronilor de mamifer sunt foarte subtiri (cei mai grosi au diametrul de 20 microni) si de aceea nu se pot inregistra variatiile de potential ale unui singur axon. Din acest motiv, s-a recurs la studiul modificarilor electrice in axonii giganti nemielinizati ai unor nevertebrate (crab, sepia etc.), care au grosimi pana la 1mm si ale caror proprietati fundamentale nu difera de cele ale axonilor de mamifer.
Potentialul membranar de repaus (PR) si potentialul de actiune (PA)
Neuronii se afla in medii apoase, iar lichidele din interiorul si exteriorul lor sunt solutii electrolitice cu un continut bogat in anioni sau cationi.
Datorita permeabilitatii selective a membranei neuronale, care nu permite trecerea libera a anionilor (cu exceptia Cl-) si a cationilor (cu exceptia K+), care par sa circule liber in ambele sensuri, ca si a unor mecanisme active "pompa" de Na+- K+) se realizeaza:
- o predominanta de anioni in interiorul celulei (anioni organici, in special fosfati, sulfati si proteine, putin Cl-, si o mare cantitate de K+, dar foarte putini Na+);
- si o predominanta de cationi in exteriorul neuronului - concentratie mare de Na+ si foarte mica de K+.
Distributia inegala de sarcini pozitive sau negative pe cele doua fete ale membranei este intretinuta prin interventia unor mecanisme de transport pasive si active, astfel incat membrana in repaus este polarizata ca un condensator, cu sarcinile negative in interior si sarcinile pozitive in exterior.
Membrana nu este insa un condensator perfect, ci are pierderi, "scurgeri de curent', care sunt mereu compensate prin transportul activ. Acesta este necesar deoarece:
- Na+ are mereu tendinta de a intra in celula, favorizat de gradientul de concentratie (concentratie de 142 mEq/1 extracelular si 10 mEq/1 intracelular) si de gradientul electric-interiorul celulei este negativ. I se opune insa membrana, care este foarte putin permeabila pt. Na+ in stare de repaus, ceea ce face ca practic difuziunea sa fie foarte mica.
- K+ are tendinta permanenta de a iesi din celula (pasiv) datorita gradientului de concentratie (150 mEq/1 intracelular si 5 mEq/1 extracelular), dar si permeabilitatii membranare, care in stare de repaus este mult mai crescuta pentru K+ (de 50-100 de ori mai permeabila decat pentru Na+).
Mentinerea diferentelor ionice de o parte si de cealalta a membranei se realizeaza prin transportul activ - pompa cuplata de Na+-K+, care functioneaza cu consum energetic, energia rezultand din scindarea ATP. Aceasta elimina 3 Na+ si introduce 1K+. .
Neuronul se poate afla in trei stari : de repaus, excitat (activat) sau inhibat.
Potentialul de repaus apare datorita repartitiei neuniforme a ionilor si deci a sarcinilor electrice, de o parte si de cealalta a membranei, care genereaza o diferenta de potential intre interiorul si exteriorul neuronului variabila intre -70 si -90 mV. Aceasta distributie neuniforma, asimetrica a ionilor se datoreaza:
- transportului activ, prin care Na+ este continuu pompat afara din celula, iar K+ este introdus;
|
- permeabilitatii membranei celulare in repaus, care pentru Na+ este mica;
- impermeabilitatii membranei pentru cei mai multi dintre anionii din interiorul celulei;
- permeabilitatii membranei pentru K+, care este relativ mare si care datorita gradientului de concentratie determina efluxul pasiv al K+;
- permeabilitatii crescute pentru Cl-, care iese din neuron.
Prin urmare, in stare de repaus, permeabilitatea mare a membranei pentru K+, cat si gradientul de concentratie favorabil permit acestuia sa iasa pasiv in exterior, unde aduce un surplus de sarcini pozitive. Efluxul se va produce pana cand sarcinile pozitive acumulate la exterior vor realiza o bariera electrostatica, care se va opune iesirii K+. In acest moment, diferenta de potential intre cele 2 fete ale membranei se stabilizeaza la valoarea de -70 sau -90 mV, adica s-a atins potentialul de repaus. Deci, PR este un potential generat de ionii de potasiu.
Pofentialul de actiune se produce atunci cand un stimul (electric, chimic, mecanic, fizic etc.), care actioneaza asupra neuronului creste brusc permeabilitatea locala a membranei pentru ioni, in general si pentru Na+ in special, modificand starea de repaus a membranei pe care o depolarizeaza. Primul eveniment in generarea potentialului de actiune il constituie cresterea brusca a permeabilitatii membranei neuronale, ca urmare a deschiderii canalelor pentru Na+, ceea ce are ca rezultat micsorarea diferentei de potential cu -15mV, care permite atingerea nivelului critic pentru declansarea potentialului de actiune. Acum se deschid canalele rapide pentru Na+, care sunt dependente de voltaj (aprox. -60;-75mV), iar Na+ va intra rapid, "in avalansa", aducand un surplus de sarcini pozitive, care vor inversa polaritatea initiala = depolarizare.
Canalele rapide sunt strajuite de doua porti h si m. In repaus poarta h este deschisa, iar poarta m este inchisa. Cand neuronul este stimulat si potentialul de repaus scade la -60 mV, adica la potentialul de prag, poarta m se deschide rapid. La un voltaj similar, aproape de prag, poarta h se inchide, insa ceva mai lent decat poarta m, astfel ca exista o scurta perioada in care ambele porti sunt complet deschise si Na+ intra rapid in neuron.
Spre sfarsitul PA se produce si o usoara crestere a conductantei pentru K+. Imediat dupa depolarizare membrana devine iarasi aproape total impermeabila pentru Na (modificarile de voltaj duc la inchiderea canaleleor de Na). Sarcinile pozitive din interiorul celulei incep sa fie eliminate prin efluxul de K, care incepe sa creasca atat datorita deschiderii canalelor pentru K+, cat si disparitiei barierei electrostatice. Interiorul va redeveni predominat negativ, exteriorul predominant pozitiv, procesul numindu-se repolarizare. Pana acum, schimbul ionic s-a realizat pasiv, favorizat de gradientul electric, ionic si de deschiderea canalelor. In acest moment, neuronul nu a revenit complet la starea de repaus, deoarece, desi s-a restabilit polaritatea de repaus, gradientul ionic este inversat, adica Na+ este in celula, iar K+ in exterior. Scoaterea Na+ din celula si introducerea K+ nu se poate face pasiv (se opun gradientele de concentratie), motiv pentru care vor intra in actiune pompele de Na+-K+. Corespondentul electric al activitatii acestor "pompe' il constituie aparitia postpotentialelor, negativ si pozitiv.
- Postpotentialul negativ pare a fi generat de cresterea K+ la exteriorul membranei, crestere care impiedica temporar restabilirea completa a PA, suprafata membranei ramanand inca depolarizata cateva minute, depolarizare reziduala.
- Postpotentialul pozitiv. In tendinta de a reveni la nivelul de repaus, potentialul devine putin mai negativ decat valorile de repaus (hiperpolarizare reziduala), ca urmare a functionarii excesive a pompei de Na+ care elimina mai mult Na+, ducand la un exces de negativitate la exteriorul celulei.
PA se insoteste de modificari ale excitabilitatii: faza refractara absoluta si faza refractara relativa. Perioada refractara absoluta face ca PA sa respecte legea "tot sau nimic', neprezentand din aceasta cauza fenomenul de sumatie.
Cea mai inalta zona de excitabilitate se afla in zona hilului axonului (denumita si pacemaker), unde pe o lungime 50-100 microni nu exista teaca de mielina. Aici pragul de excitabilitate este de aproximativ trei ori mai mic decat in alte zone ale neuronului, ceea ce face ca de aici sa inceapa depolarizarea neuronului, propagandu-se apoi in soma neuronala.
Propagarea impulsului nervos
In cazul fibrelor amielinice propagarea impulsului se face din aproape in aproape prin curenti de depolarizare - curentii locali Hermann. Procesul consta in depolarizari si repolarizari succesive, intre segmentul stimulat (negativ la exterior) al axolemei si zonele invecinate, inca pozitive aparand o diferenta de potential, care genereaza un curent electric local (de la pozitiv la negativ), cu o durata de cateva milisecunde Acest curent are o tensiune suficient de mare pentru a genera in regiunea invecinata un potential de actiune care se propaga din aproape in aproape, ca "praful de pusca aprins" sau ca "focul in preerie. Desi se propaga decremential (cu pierdere de energie electrica in mediul extraaxonal), datorita rezistentei mici a celulelor Schwan si a faptului ca PA are o valoare mult mai mare decat pragul de stimulare (de 5-10 ori mai mare), PA se poate propaga pana la celula urmatoare. Acest tip de propagare se face cu o mare cheltuiala de energie si o viteza foarte redusa, fiind caracteristic fibrelor vegetative si somatice subtiri (sub 1 micron)
In fibrele mielinice, al caror segment internodal este practic inexcitabil (avand o rezistenta mult mai mare decat la nivelul nodului si fiind impermeabil pentru ioni) biocurentii generati in timpul excitatiei strabat membrana numai la nivelul nodurilor Ranvier, propagarea potentialului facandu-se in salturi. Depolarizarea inainteaza de-a lungul fibrei, fiind urmata, cu o intarziere de cateva zeci de milisecunde, de procesul de repolarizare, care se raspandeste progresiv de-a lungul fibrei, in aceeasi directie ca si depolarizarea, restabilind starea initiala a fibrei. Conducerea "saltatorie" creste viteza conducerii impulsului de 10-50 ori comparativ cu aceea din fibrele amielinice, iar consumul energetic este de 300 de ori mai mic.
Frecventa maxima a PA transmise printr-o fibra nervoasa este limitata de perioada refractara (1 ms.), fiind teoretic estimata la 1000 impulsuri/s., valoare rar atinsa, deoarece frecventa obisnuita este cuprinsa intre 5-100 impulsuri/s. si doar rareori depaseste 200 impulsuri/s.
Proprietatea nervului de a conduce unde de depolarizare se numeste conductibilitate si are la baza o serie de legi :
- legea integritatii fiziologice a fibrei nervoase. Conducerea impulsului nervos necesita integritatea anatomica si functionala a nervului, un nerv comprimat, ligaturat, tractionat sau novocainizat nu mai conduce impulsul;
- legea conducerii izolate. Fiecare fibra nervoasa conduce numai impulsurile proprii, fara ca acestea sa se transmita si fibrelor invecinate din acelasi trunchi nervos (astfel se efectueaza miscari precis coordonate). Conducerea izolata este posibila datorita prezentei tecii de mielina, dovada fiind ca in axonii partial lezati excitatia difuzeaza, se transmite si fibrelor paralele invecinate (conducere ephaptica).
- legea conducerii bilaterale se refera la capacitatea fibrelor nervoase de a conduce excitatia in ambele sensuri, centripet si centrifug, capacitate limitata insa de prezenta sinapsei, unde excitatia se propaga intr-un singur sens ortodromic, de la componemta presinaptica la cea postsinaptica.
- legea conducerii nedecrementiale - impulsul nervos este condus de-a lungul fibrei nervoase fara nici o pierdere, amplitudinea undei mentinandu-se aceeasi de-a lungul fibrei, indiferent de lungimea acesteia.
- legea multiplicarii impulsului nervos la nivelul terminatiilor axonale - un singur neuron motor poate stabili prin butonii terminali contacte (sinapse) cu mai multi neuroni, carora le transmite impulsul nervos nedecremential.
- legea "tot sau nimic"- fibrele raspund maximal la un stimul prag sau supraprag, iar odata generat un potential de actiune in oricare punct al membranei, acesta va fi transmis nedecremential pe intreaga membrana.
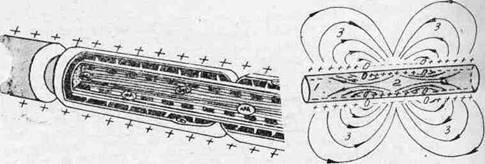
Propagarea impulsului in fibrele amielinice.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Analize pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||